![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ЦИНК
Первое упоминание о цинке, как о микроэлементе, влияющем на рост растений, относится к 1869 г., когда Дж. Ролен, добавив ничтожное количество этого элемента к грибкам и водорослям, обнаружил положительное влияние цинка на развитие низших растений. Необходимость цинка для высших растений была показана в 1872 г. нашим соотечественником К. А. Тимирязевым. Сведения по содержанию цинка в растениях были собраны многими исследователями в первой трети XX столетия. В 1914 г. показано, что кукуруза не может нормально расти без цинка. Через пять лет, в 1919 г., отсутствие цинка в питательной среде вызвало значительную (почти в 2000 раз) потерю массы мицеллия гриба. Систематические сведения о значении цинка для нормального роста и развития растений появились в печати в конце 20-х и начале 30-х годов. В 1926 г. обнаружилось, что ячмень и подсолнечник вовсе не могут развиваться без цинка. В 1928 г. опыты с гречихой и бобами показали необходимость цинка для нормального их роста и развития. У бобов отсутствие цинка вызывало опадение листьев и цветочных почек. В 1954 г., выращивая 26 различных культур без добавления цинка и с внесением удобрений, установили, что одни растения в большей, а другие в меньшей степени нуждаются в цинке. Для сравнительной оценки действия цинка к настоящему времени систематизированы результаты анализа большого количества растений. Теперь установлено, что цинк входит в состав различных органов всех растительных организмов, но содержание этого элемента в различных растениях очень разнообразно.
Способность извлекать цинк из почвы у разных видов растений далеко не одинакова. Например, люцерна находила для себя этот элемент там, где злаки страдали от цинковой недостаточности. Присутствие люцерны в садах предупреждало появление мелколистно-розеточной болезни, вызываемой недостатком цинка у плодовых деревьев. Сорные растения вообще содержат больше цинка, чем культурные. Наиболее высокое количество цинка у ядовитых грибов; много этого элемента в лишайниках и хвойных деревьях. Не в этом ли объяснение бурного роста различных сорняков?
Содержание цинка в растениях и вынос из почвы урожаями различных культур на черноземе (по данным М. В. Каталымова)
|
Название растения |
Содержание цинка растениями, мг/кг сухого вещества |
Вынос цинка, г/га |
Название растения |
Содержание цинка растениями, мг/кг сухого вещества |
Вынос цинка, г/га |
|
Фасоль: |
|
|
Овес: |
|
|
|
семена . . . |
33,0 |
54 |
зерно . . . . |
36,0 |
|
|
солома , , . |
20,0 |
|
солома . . . |
35,0 |
145 |
|
Люцерна: |
|
|
Свекла кормовая |
|
|
|
. сено . . . . |
25,0 |
75 |
корни . . . |
25,0 |
153 |
|
Пшеница яровая: |
|
|
.листья . |
22,5 |
|
|
зерно . . . . |
65,0 |
87 |
Подсолнечник: |
|
|
|
солома , , . |
16,0 |
|
семена . . . |
52,5 |
174 |
|
Ячмень: |
|
|
стебли, листья |
25,0 |
|
|
зерно . . . . |
38,0 |
97 |
Свекла сахарная |
|
|
|
солома . . . |
20,0 |
|
. корни - . . . |
17,5 |
188 |
|
|
|
|
листья . . . |
50,0 |
|
Многочисленными определениями показано, что количество цинка в растениях колеблется от десятитысячных до сотых долей процента. У различных растений, выращенных в одних и тех же условиях (например, на черноземе)-, содержание цинка составляет от 18 до 65 мг на 1 кг сухого вещества, В этом случае изменение количества цинка в растениях связано с их биологической особенностью. Неодинаков и вынос цинка урожаями сельскохозяйственных культур (табл. 17). Здесь нет прямой пропорциональной зависимости, но картина получается довольно наглядной. Больше всего собирают цинка из почвы с урожаями свеклы, подсолнечника, проса и овса, меньше всего— с урожаями фасоли, люцерны и пшеницы. Минимальное и максимальное количества цинка, выносимого из почвы урожаями сельскохозяйственных культур, отличаются друг от друга более чем в 3 раза и составляют 54 и 188 г на 1 га пахотной земли. ,
Поступление цинка в растения и вынос его с урожаем сельскохозяйственных культур определяются не только -биологической особенностью отдельных растений, но и свойствами почв, на которых они выращиваются. Если сравнить содержание цинка в растениях и вынос его с урожаем одних и тех же культур, возделанных на мощном черноземе и на дерново-подзолистых почвах, то нетрудно заметить, что на дерново-подзолистом суглинке в растения поступает и выносится с урожаем больше цинка, чем на мощном черноземе. Эта простая, но очень важная зависимость для жизнедеятельности растений была проверена на главных почвенных типах нашей страны применительно к четырем культурам: ячменю, льну, красному клеверу и горчице.
В зависимости от типа почвы поступление цинка в растения существенно изменялось: для ячменя в 2 раза (от 40 до 80 мг), для льна (от 38 до 115 мг) и клевера (от 60 до 180 мг) в 3 раза, для горчицы даже более чем в 4 раза (от 106 до 437 мг). Больше всего цинка из почвы поступает к горчице, меньше всего — к ячменю и льну, красный клевер занимает среднее положение» Из приведенных данных выявляется одна интересная и важная особенность для сельского хозяйства. Известкование почвы снижает поступление цинка в растения; наибольшее количество цинка извлекается из неизвесткованной дерново-подзолистой почвы. Во всех других случаях усвоение цинка растениями было заметно ниже. Опыты со льном показали, что увеличение дозы извести» вводимой в почву, снижает содержание цинка в растениях (данные М. В.'Каталымова):
Дозы извести, т/га 0 2,25 4,6 9,0 13,5 22,5
Содержание цинка у льна, мг/кг 55,0 51,3 49,0 48,3 46,7 36,3
Для различных органов растений была установлена одна общая закономерность: больше всего цинка содержится там, где много хлорофилла. В листьях овса найдено 20—30% цинка от общего количества его в растениях. Повышенное содержание цинка обнаружилось в листьях пшеницы (по сравнению с ее стеблем). Более высокое сосредоточение цинка отмечено в точках роста растения, в его генеративных органах. Ученые показали, что цинк в больших количествах поступает в молодые растущие органы растений. Однако наибольшее скопление цинка (так же как бора и меди) отмечено в зародышах семян. Эти факты свидетельствуют, что с цинком связаны важнейшие процессы жизнедеятельности растений, в том числе такие, как фотосинтез и размножение. Есть основания полагать, что цинк в процессах размножения играет общебиологическую роль.
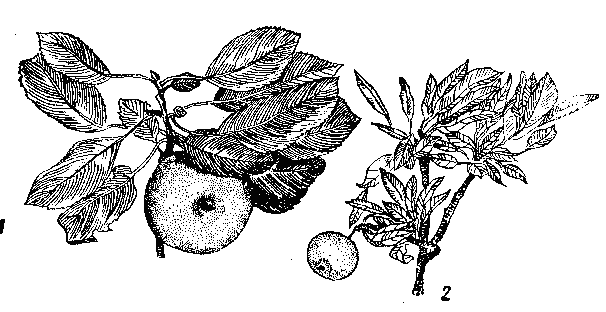
Рис. 28. Недостаток цинка у яблони:
1 — листья и плод» получившие цинк; 2 — узкие деформированные хлоротичныв листья и мелкий плод при недостатка цинка.
Недостаток цинка, а точнее, его подвижной формы в питательной среде приводит к патологическим изменениям в растениях. Среди них главным является задержка роста, приводящая к почти полному прекращению роста междоузлий. И как следствие этого — появление приземистых форм растений. Такое ненормальное развитие растений объясняется содержанием ауксина, причем уменьшение цинка в растениях приводит к понижению в них ауксина. Причиной уменьшения ауксина при цинковой недостаточности является, по-видимому, не столько усиление его окисления, сколько ухудшение биосинтеза -предшественников ауксина, например индолилукеусной кислоты, триптофана и других аминокислот. Другим симптомом цинковой недостаточности, в частности, у листопадных плодовых деревьев, таких, как яблони, является небольшой размер листьев, составляющий от нормальной их величины,- располагающихся на ветке в форме розетки. Это заболевание называется мелколистностью и розеточностью (рис. 28). На деревьях, страдающих такой болезнью плодовых почек образуется мало, а появляющиеся немногочисленные плоды бывают - небольшой величины и уродливой формы. На листьях больших растений, приобретающих ненормальную форму, появляются хлоротические пятна, которые становятся светло-зелеными, а иногда желтыми и даже белыми. Но мере развития болезни у деревьев через несколько лет начинают отмирать ветви, а при более сильном цинковом голодании возможна гибель растении, Кроме яблонь, мелколнстностью и розеточностью могут заболеть и другие плодовые деревья: груша, слива, вишня, черешня, виноград, персики, абрикосы (рис, 29).
От недостатка цинка заболевают и цитрусовые культуры: мандариновые, апельсиновые и лимонные деревья. Болезнь эта носит название «пятнистость» или «крапчатость» листьев и во многом напоминает мелколистность и розеточность яблонь. На листьях цитрусовых деревьев появляется хлоротическая крапчатость листьев.
Возникает она вначале на самых нижних побегах и постепенно распространяется R вершине дерева. Листья между жилками становятся желто-зелеными, а по мере их старения — багрово-красными. Такие листья начинают преждевременно опадать, вызывая оголение побегов, на концах которых образуются розетки из мелких листьев. При значительном цинковом голодании эти листья становятся очень твердыми. Если на таких больных деревьях появляются плоды, то бывает их очень мало, они имеют неправильную форму и обладают плохими вкусовыми качествами. После 8—4 лет цинкового голодания деревья погибают. Отмечена болезнь цинковой недостаточности у грецкого ореха (желтуха) и у тунгового дерева (бронзовость листьев).
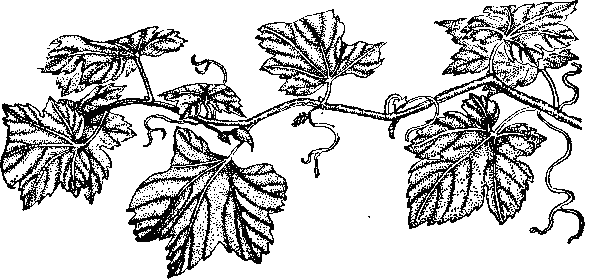
Рис. 29. Недостаток цинка , у винограда: межжилковый хлороз
листьев. .
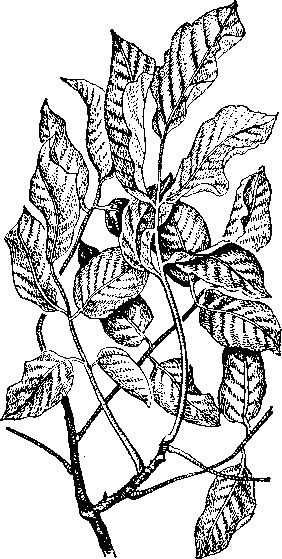
Рис. 30. Недостаток цинка у грецкого ореха: слабый хлороз.
Симптомы за болевания практически те же самые.
У грецкого ореха образуются мелкие листья, располагающиеся в форме розетки (рис. 30); у тунгового дерева заболевание обнаруживается в мае —июне по образованию бронзовой окраски листьев, которая затем темнеет и переходит в некротические пятна отмершей ткани листья становятся как бы разорванными.
Из травянистых растений цинковая недостаточность чаще всего проявляется у кукурузы. К внешним признакам болезни относятся такие признаки: побеление верхушки, образование желтых хлоротнческих полос между жилками листьев, появление светло-желтой, иногда даже белой окраски у вновь образующихся листьев, отмирание нижних листьев.
Известны, хотя и достаточно редко встречаются в полевых условиях, также общие признаки цинковой недостаточности у овощных культур: картофеля, томатов, тыквы, горчицы (рис. 31). Прежде всего это пятнистость, хлороз, уменьшение листовой пластинки. У бобовых симптомы цинкового голодания проявляются при содержании цинка в листьях менее 0,002%. Однако болезнь не дает о себе знать вплоть до цветения этого растения. В период цветения бобовые быстро теряют листья, к тому же стручков, не содержащих семян образуется мало. Из бобовых культур фасоль и соя являются наиболее чувствительными к болезни, вызванной отсутствием цинка.
Все растения, не исключая плодовые деревья подвержены болезни цинковой недостаточности. В зимнее время очень чувствительны к морозам, неустойчивы против них и постепенно засыхают. Давно уже подмечено, что. количество цинка в растениях в какой-то степени зависит от метеорологических условий. В те годы, когда выпадает большое количество осадков, содержание цинка в растениях увеличивается по сравнению с годами, когда осадков бывает поменьше.
Физиологическая роль цинка — его многостороннее влияние на жизнь и развитие растений — изучалась многими исследователями. В конце 30-х и начале 40-х годов были проведены исследования, показавшие структурные изменения в клетках при цинковом голодании и сыгравшие определенную роль в понимании физиологического значения цинка. Этот микроэлемент принимает участие в деятельности металлоферментов и активизации металлоферментных комплексов. В настоящее время из- 800 известных ферментов около 30 относится к цинксодержащим, число которых все время увеличивается (достаточно сказать, что в 1952 г.- таких цинксодержащих ферментов было только 7).
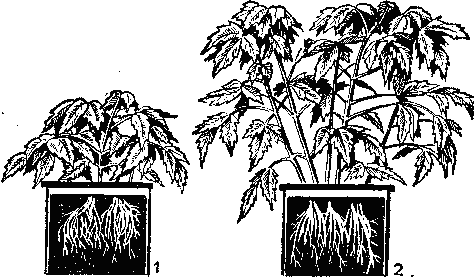
Рис. 31. Влияние цинка на рост томатов:
1— без цинка; 2 — с цинком.
Первым ферментом, где в ,1932 г. был обнаружен цинк, является карбоангидраза. Этот фермент полученный в виде кристалла представляет собой сложное образование, состоящее из белка и небелкового компонента, называемого простетической (активной) группой. Отличительной особенностью последней является то, что она по существу определяет каталитические способности фермента. Белковая же часть фермента обусловливает специфические свойства их избирательность действия на строго определенный процесс. Содержание цинка в карбоангидразе составляет 0,33—0,34%, он прочно связан в белковой молекуле этого фермента.
Нахождение цинка в карбоангидразе у растений было установлено в 1946 г. Этот фермент оказался катализатором обратимой реакции расщепления угольной кислоты на оксид углерода (IV) и воду.
Было показано что цинк в составе рассматриваемого фермента участвует в выделении углекислого газа. Карбоангидраза, способствуя улавливанию его, оказывает влияние на дыхание растений. Можно определенно утверждать, что снижение активности цинксодержащих ферментов безусловно сказывается на работе других самых разнообразных , ферментативных систем и может вызвать отклонения в реакциях -дыхания растений. Для всех культур, содержащих карбоангидразу (например, овес, горох, петрушка, томаты), установлена определенная зависимость между уменьшением или увеличением количества цинка и активностью карбоангидразы в растениях; активность карбоангидразы снижается при недостатке цинка.
Цинк является компонентом очень многих дегидрогеназ. Наиболее изученной из всех известных дегидрогеназ является алкогольдегидрогеназа, в которой на одну молекулу белка приходится 4 атома цинка. Алкогольдегидрогеназа играет определенную роль в образовании пигментного аппарата растений, а недостаток цинка в листьях растений снижает ее активность. Это приводит к уменьшению размеров пластид и образованию в них вакуолей, содержащих фенольные соединения, а также к сильному поражению хлоропластов, которые, сливаясь и собираясь в группы, задерживаются в своем развитии или совсем разрушаются. Алкогольдегидрогеназа содержится в пшенице, горохе, кукурузе, рисе, сахарной свекле, петрушке, шпинате, табаке, листьях мяты перечной, семенах многих -растений в грибах и бактериях. При цинковой недостаточности в грибах отмечено ослабление интенсивности окислительных процессов и накопление промежуточных, менее окисленных соединений. В этом случае наблюдается отсутствие фермента алкогодьдегидрогеназы, обычно обнаруживаемого в грибах.
Механизм активирования цинком кристаллической дрожжевой алкогольдегидрогеназы (сокращенно ДАД) в присутствии никотинамидадениннуклеотида (НАД), по данным Хоха и Валл и, представляется так:
Из схемы видно, что цинк связывает с возникновением ферменткоэнзимного комплекса. Диссоциация этого комплекса определяет протекание всех остальных реакций. Цинк, обеспечивая каталитическую функцию фермента, служит и для ее стабилизации.
Цинк входит в состав целого ряда ферментов и активирует деятельность многих из них, например щавелеуксусную дегидрогеназу, ответственную за образование пировиноградной кислоты и отщепление С02 от щавелеуксусной кислоты. Поскольку пировиноградная кислота является основным звеном обмена веществ в растительной клетке, то можно полагать, что цинк влияет на образование пировиноградной кислоты и поэтому имеет важное значение для процессов обмена в растительных организмах. Другой цинксодержащий фермент —триозефосфатдегидрогеназа участвует в окислении и фосфорилировании фосфогдицеринового альдегида с образованием дифосфоглицериновой кислоты и регулирует поступление фосфора в растения. В условиях цинковой недостаточности - активность триозефосфатдегидрогеназы снижается и начинается повышенное поступление неорганического фосфора в листья, стебли и корни растений. Одновременно уменьшается количество различных органических соединений фосфора (общего фосфора, нуклеотидов, лабильного фосфора, фосфора липидов и нуклеиновых кислот) в растениях. Например, цинковая недостаточность приводит к резкому увеличению содержания неорганического фосфора в ячмене, горохе, томатах и снижению содержания фосфора, входящего в состав нуклеотидов, липоидов и нуклеиновых кислот. Нарушения в поглощениях фосфора растениями нормализуются после добавления цинка в питательную среду. Значительна роль цинка в окислительно-восстановительных процессах, протекающих в растениях. При ухудшении условий питанием цинком наблюдаются функциональные расстройства и нарушение окислительно-восстановительных систем. Этот микроэлемент в подобных процессах по сравнению с марганцем и медью играет несколько иную роль. Марганец и медь усиливают окислительные процессы с образованием в растениях более окисленных соединений, а цинк в общей цепи окислительно-восстановительных процессов способствует усилению прямо противоположных процессов — восстановительных.
Нарушение окислительно-восстановительных процессов приводит к нарушению синтеза белка в растениях. Основной причиной этого является снижение активности различных ферментов в отсутствие или при недостаточности в них цинка. У томатов и овса отмечено снижение белка и значительное накопление небелковых растворимых соединений азота: почти двойное увеличение количества аминокислот и десятикратное увеличение амидов. Известно, что аминокислоты представляют собой как бы осколки белковой молекулы, подвергнувшейся гидролитическому расщеплению под воздействием некоторых ферментов. Амиды в свою очередь получаются из аминокислот путем замещения гидроксила кислоты аминогруппой.
Есть основания полагать, что, оказывая влияние на нуклеиновые кислоты, цинк (необходимый, как теперь выяснилось, для их получения) регулирует синтез белка. Подтверждением этого является снижение содержания рибонуклеиновой кислоты РНК при недостатке цинка в листьях цитрусовых деревьев, яблонь, винограда, у томатов, ячменя, кукурузы.
Участие цинка в окислительно-восстановительных реакциях и регулировании активности ферментов синтеза белка определяет влияние цинка на фотосинтез, образование хлорофилла и рост растений. Еще в 1940 г. было показано, что содержание хлорофилла у одноклеточной водоросли хлореллы зависит от наличия цинка. Отсутствие цинка приводило к уменьшению содержания хлорофилла в ней. Внесение этого элемента помогало очень быстрому восстановлению прежнего нормального содержания хлорофилла. Не исключена возможность, что с этим процессом связаны пигментные изменения, приводящие к различному содержанию хлоропластов у растений.
Таким образом, значение цинка для физиологии растений объясняется широким кругом процессов, в которых он принимает участие. Прежде всего в фотохимических реакциях. Как часть ферментов углеводного обмена (альдолазы, энолазы, карбоксилазы и др.) он активирует их и способствует повышению интенсивности фотосинтеза. При недостатке цинка в листьях растений наблюдается скопление редуцирующих сахаров и в меньших количествах сахарозы и крахмала. Цинковая недостаточность нарушает обмен крахмала и приводит к расстройству углеводного обмена. Можно отметить влияние микроэлемента на синтез и гидролиз углеводов. Выяснилось, что форма азота, которую получают растения, оказывается при этом решающим фактором. Если растения получают азотное питание в аммиачной форме, то количество сахарозы, например, в листьях гречихи и пшеницы увеличивается, а содержание моносахаров уменьшается. При нитратном питании растений цинк, уменьшая поступление сахаров в эти растения, в то же время увеличивает поступление в них моносахаров. И наконец, недостаток цинка сильно отражается на растениях с высоким содержанием углеводов.
Во время засухи цинк, увеличивая количество фосфатидов и нуклеопротеидов в растениях, нормализует их энергетические процессы. Поскольку фосфатиды являются посредниками в синтезе жирных кислот, то всякое возрастание количества фосфатидов приводит к увеличению количества в клетках растений жиров и жироподобных веществ, от которых и зависит засухоустойчивость растений. Особенно велика роль цинка, если этот элемент вносить под растения в критические для них периоды в отношении водоснабжения. Роль цинка в повышении стойкости растений к различным неблагоприятным условиям объясняется способностью этого элемента сохранять наиболее высокое количество нуклеиновых кислот при самых минимальных возможностях их синтеза. Эта особенность цинка имеет большое практическое значение для сельскохозяйственных культур, выращиваемых в условиях Сибири и Дальнего Востока.
Суммируя данные о физиологической роли цинка в жизнедеятельности растений, можно вполне определенно говорить о многофункциональности этого элемента, так как недостаток цинка в питании растений приводит к нарушению их важнейших физиологических процессов.
![]()
![]()
![]()
![]()